原名:Invited review: Mineral absorption mechanisms, mineral interactions that affect acid–base and antioxidant status, and diet considerations to improve mineral status.
译名:特邀综述:矿物质吸收机制,影响酸碱和抗氧化状态的矿物质相互作用,以及改善矿物质状态的饲粮因素
作者:Jesse P. Goff1
完成单位:
Biomedical Sciences, College of Veterinary Medicine, Iowa State University, Ames 50011.

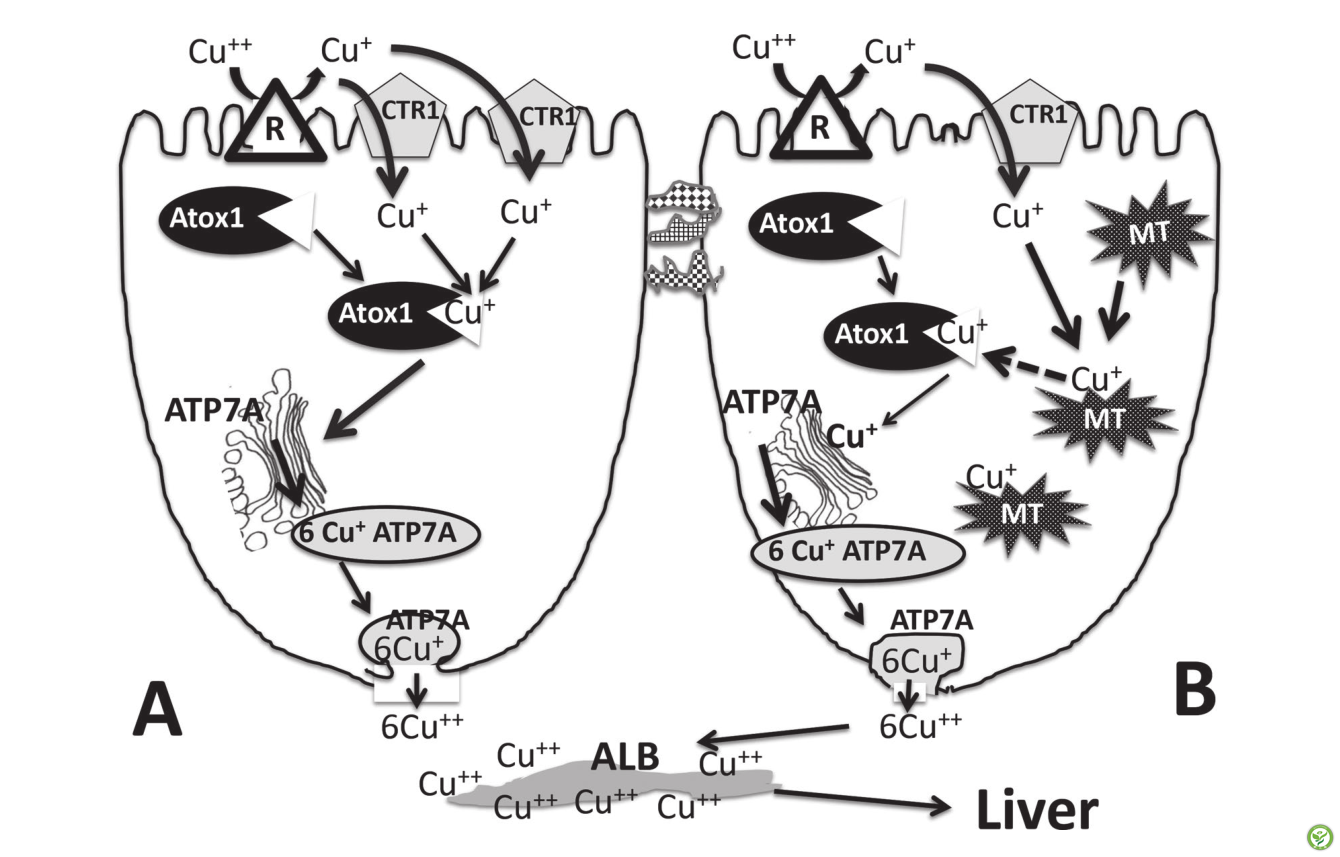
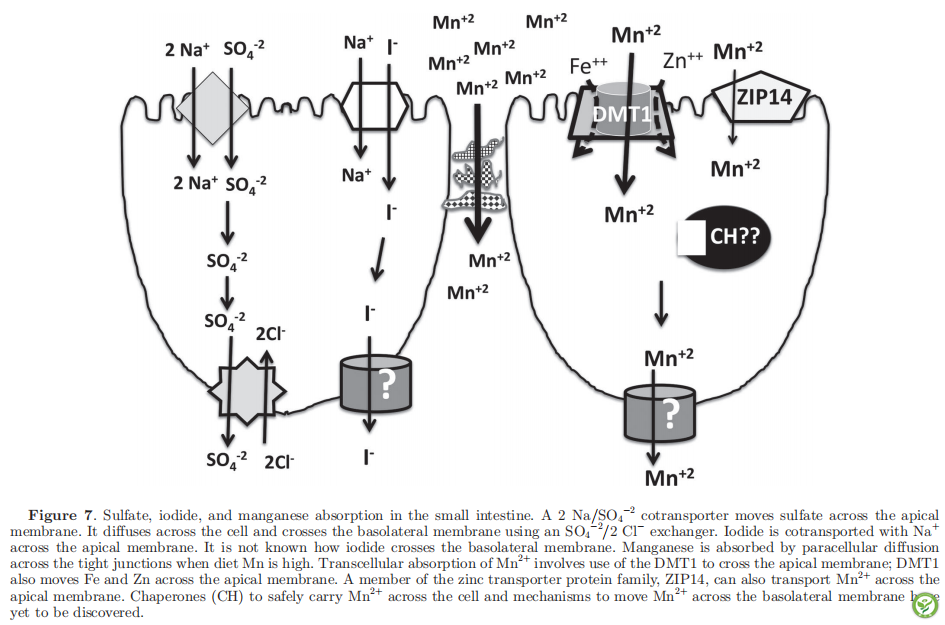
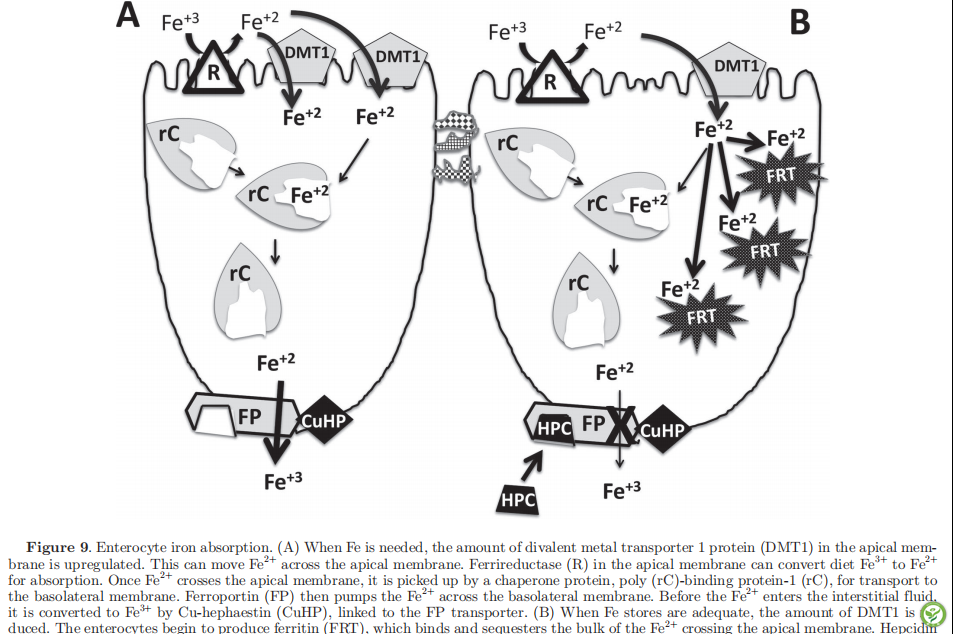
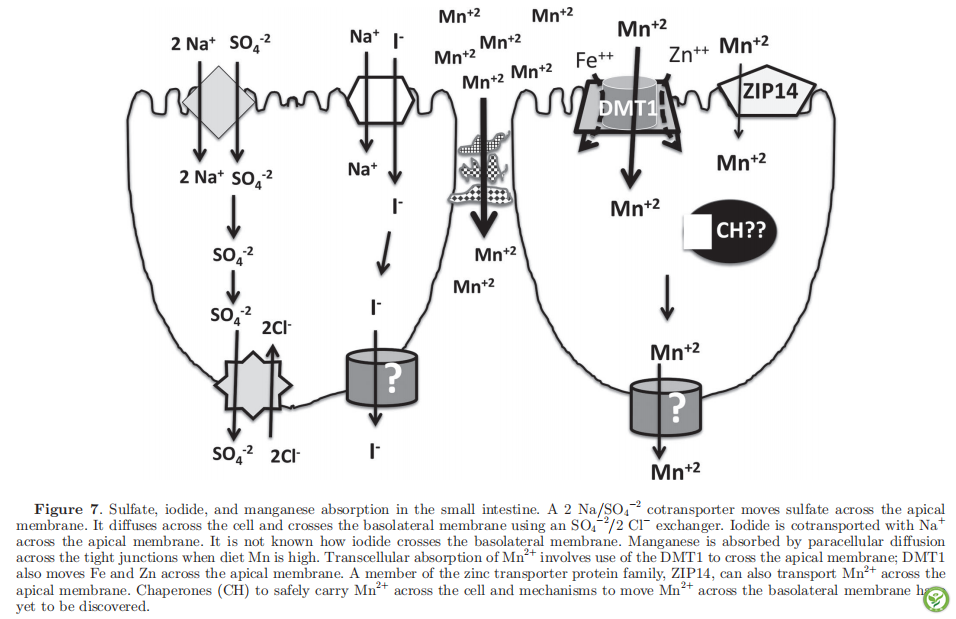
 电话: 010-80854808
电话: 010-80854808
 传真:010-89525360
传真:010-89525360
 邮箱:dysbio@163.com
邮箱:dysbio@163.com
