原标题:肠道菌群中产丁酸梭状芽孢杆菌的减少促使肠腔内需氧型沙门氏菌的繁殖
Depletion of Butyrate-Producing Clostridia from the Gut Microbiota Drives an Aerobic Luminal Expansion of Salmonella
作者:Fabian Rivera-Cha´ vez,1 Lillian F. Zhang,1 Franziska Faber,1 Christopher A. Lopez,1 Mariana X. Byndloss,1 Erin E. Olsan,1
Gege Xu,2 Eric M. Velazquez,1 Carlito B. Lebrilla,2 Sebastian E. Winter,3 and Andreas J. Ba¨ umler1,*
1Department of Medical Microbiology and Immunology, School of Medicine, University of California at Davis, One Shields Avenue, Davis,CA 95616, USA
2Department of Chemistry, College of Letters and Sciences, University of California at Davis, One Shields Avenue, Davis, CA 95616, USA
3Department of Microbiology, University of Texas Southwestern Medical Center, 5323 Harry Hines Boulevard, Dallas, TX 75390, USA
来源:Cell Host & Microbe 19, 443–454,doi:10.1016/j.chom.2016.03.004
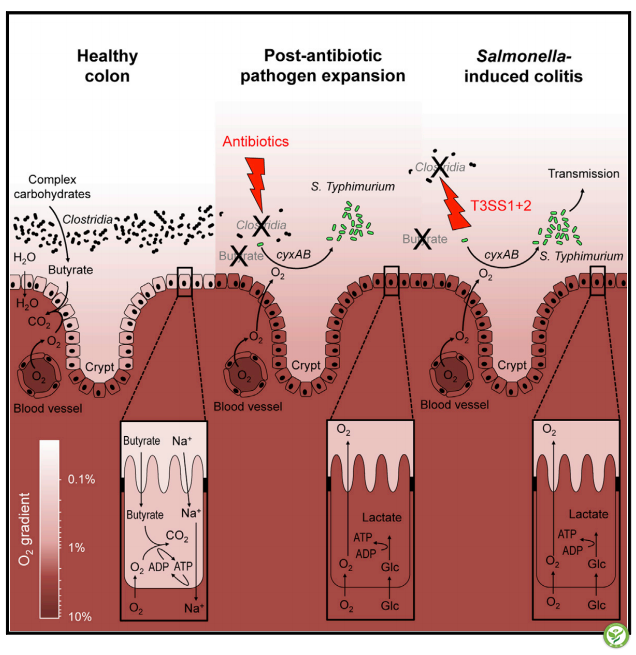
【概要】抗生素处理可促进沙门氏菌性胃肠炎复发。本文揭示:抗生素处理导致丁酸产生梭菌的减少,从而降低了沙门氏菌的定植抗性。丁酸水平的降低促进上皮细胞氧化,因而促进肠腔内需氧病原菌增殖。需氧呼吸协同硝酸盐的呼吸作用促进沙门氏菌的粪口传播。
【要点】:
沙门氏菌引发的结肠炎导致丁酸产生梭菌的减少
抗生素介导引发的梭菌减少促进结肠上皮细胞氧化
上皮细胞氧化的增加促进抗生素处理后的需氧病原菌的增殖
肠道内的由呼吸作用引发的沙门氏菌增殖是沙门氏菌粪口传播必要条件
翻译:肠动力研究院 梁琦
【摘要】哺乳动物的肠道是肠道菌群的寄生场所,其通过一些未知机制阻止病原体增殖,使用抗生素容易增加动物对肠道病原体的易感性。本试验研究了经链霉素处理的小鼠可减少肠腔中产丁酸梭状芽孢杆菌的数量,从而降低肠道内丁酸浓度,促进上皮细胞氧化,促进需氧鼠伤寒沙门氏菌的繁殖。研究发现:三丁酸甘油酯处理可恢复小鼠肠道上皮细胞低氧浓度以及抑制鼠伤寒沙门氏菌的繁殖。在无链霉素处理组,有遗传抗性的小鼠中也观察到梭状芽孢杆菌的减少和需氧型沙门氏菌的扩增,但其动力学进展较慢并且需要功能性沙门氏菌III型分泌系统的存在。沙门氏菌的细胞色素bd-II氧化酶与硝酸盐还原酶可协同促进小鼠沙门氏菌肠腔内的扩张,两者都是粪口传播的必要条件。研究结果表明:沙门氏菌毒力因子和抗生素促进病原体扩增的机制相同,即:通过减少产丁酸梭状芽胞杆菌的数量,从而促进上皮细胞氧化,进而促进需氧沙门氏菌的繁殖。
以下是实验中相关图表
图1:细胞色素bd-II氧化酶有助于抗生素处理后病原体的扩增
(A和B)用指定菌株接种培养基,并在8%,0.8%或0%氧气浓度下接种24小时后测定竞争指数(CI)
C.用指定的菌株混合物腹腔内注射感染C57BL / 6小鼠组(n = 4),感染后4天测定CI
A.感染后4天测定结肠内容物中的CI
B.感染后2天,用17种人源性梭菌分离物(C17)的培养物对一些小鼠进行胃内接种在感染前4小时和感染后2天对小鼠补充三丁酸甘油酯(TB),并在感染4天后测定CI。对于E和L,在感染一天后,部分小鼠灌胃给予氯仿处理的初治小鼠盲肠内容物(CTCC)
C.在用鼠伤寒沙门氏菌菌株感染小鼠4天后从结肠内容物中回收菌种滴度。 对于(F)和(H),每个圆圈代表来自每个动物的数据
D.在相应的时间点测定存在于20ng总细菌DNA中基于梭菌16S rRNA的基因复制数。
E.链霉素处理后的指定时间点测量盲肠内容物的丁酸浓度(参见图S1B和S1C)
F.测定存在于20ng总细菌DNA中的梭菌16S rRNA基因复制数
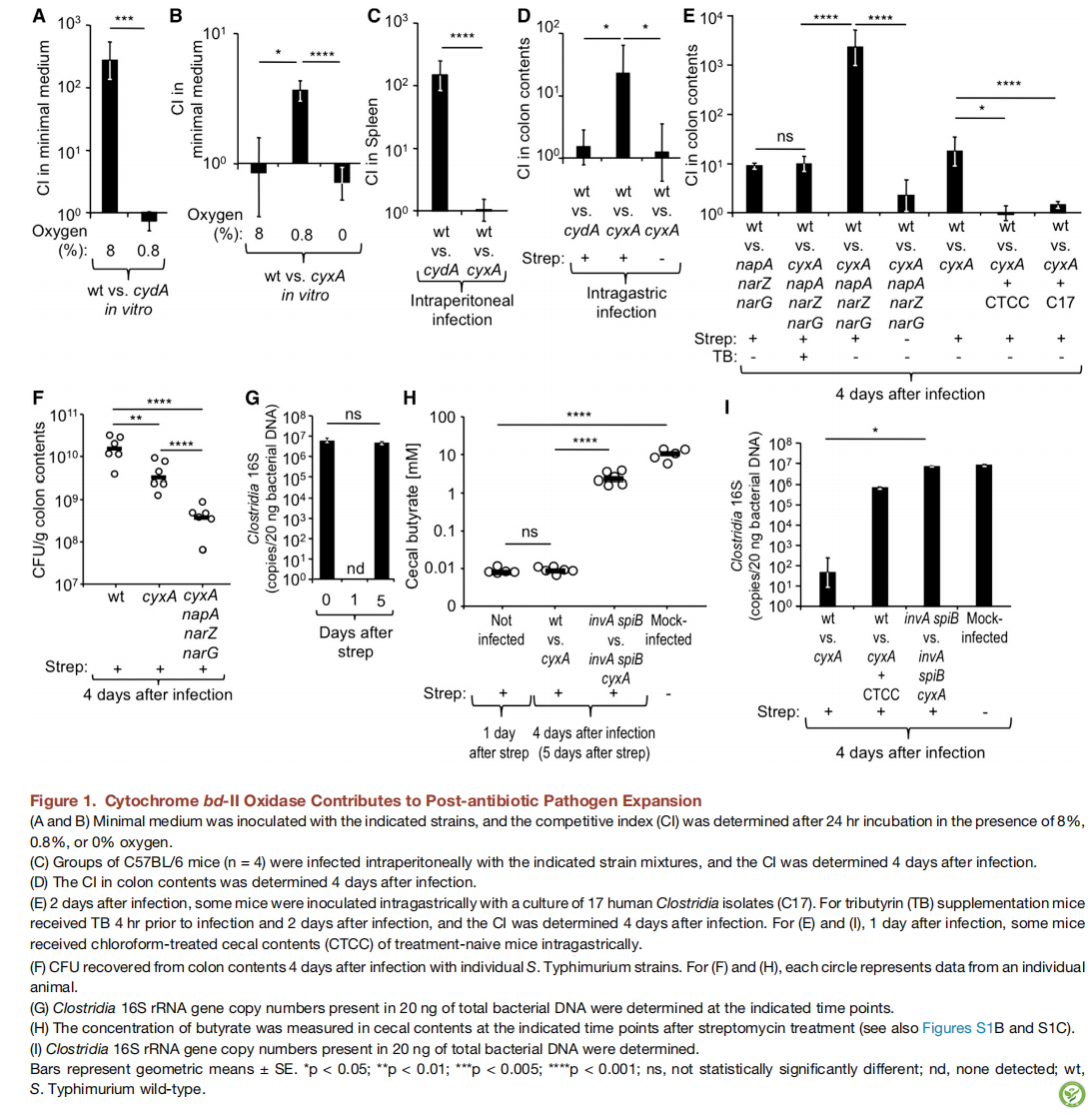
图2:三丁酸甘油酯治疗恢复结肠细胞生理缺氧并防止依赖细胞色素bd-II氧化酶的抗生素处理后的病原体扩增
A.C图)对C57BL / 6小鼠(n = 4)进行模拟处理(A),用链霉素(Strep)处理(B),或用链霉素和三丁酸甘油酯(TB)处理(C),1天后收集结肠。在用DAPI核染色剂(蓝色荧光)复染的结肠切片中检测到哌莫硝唑(红色荧光)的结合。代表性的图像于下面显示。
(D-F图)分别为模拟,链霉素处理,或用链霉素和TB处理的C57BL / 6小鼠(n = 5),在补充TB后8小时测定小鼠盲肠内容物中丁酸(D),丙酸(E)和乙酸(F)的浓度。(G和H)C57BL / 6小鼠(n在图S1A中显示,除了TB组,其余n = 4)模拟处理或用链霉素和1天后用所示的菌株混合物感染处理。一些小鼠模拟处理或在注射哌莫硝胺之前8小时补充三丁酸甘油酯,感染后1天,测定结肠内容物中的竞争指数(CI)。
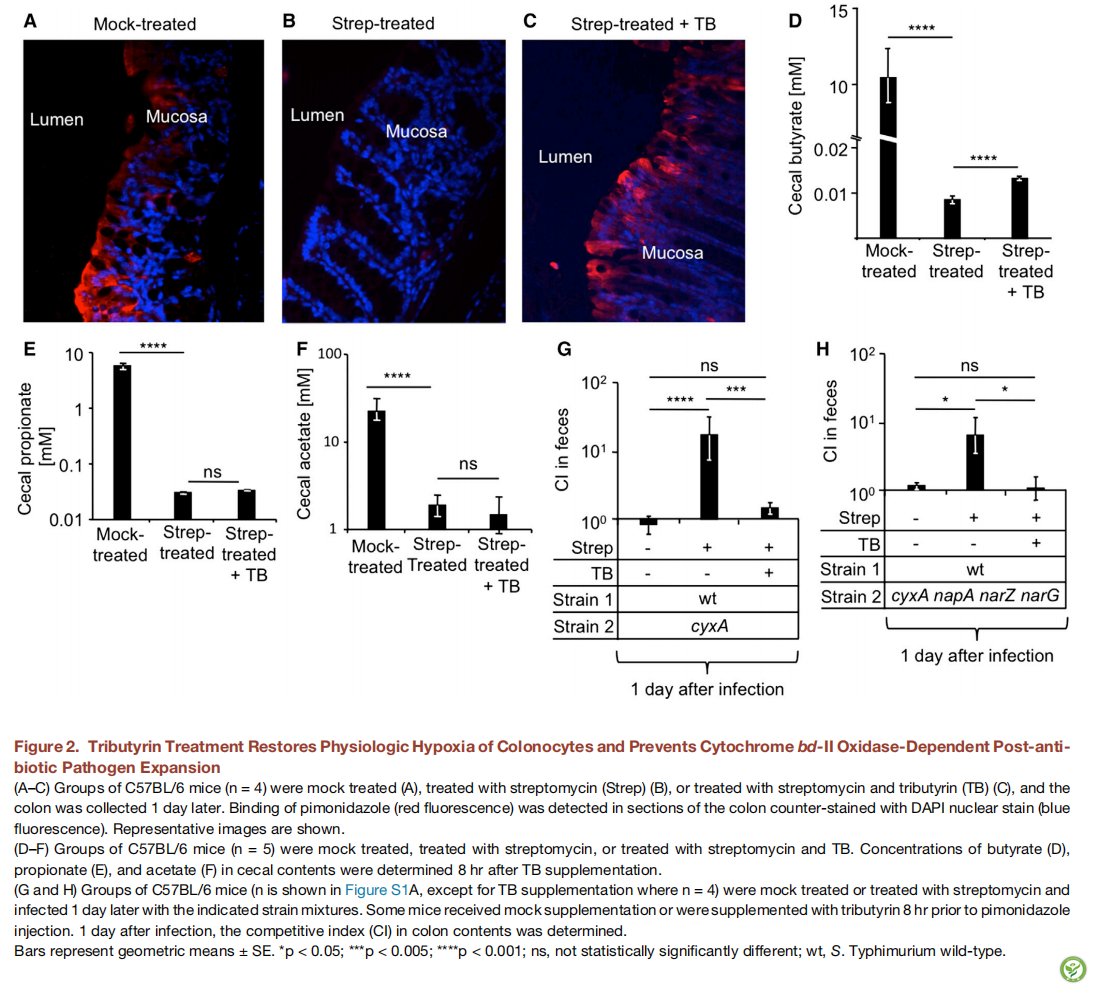
图3:毒力因子在感染后的相应时间点驱动依赖细胞色素bd-II氧化酶的鼠伤寒沙门氏菌扩增。(A-C图)C57BL / 6小鼠(n在图S1A中显示)模拟处理或用链霉素和1天后用所示的菌株混合物感染处理。(A图)在感染后第1天和第4天(第p.i.天),测定结肠内容物中的竞争指数(CI)。(B图)使用表S1中列出的标准,每组四只小鼠感染后4天,对收集的组织进行盲肠组织病理学评分(也参见图S2)。每个条形代表一只动物的数据。(C图)在(B图)中参与评分的H&E染色的盲肠切片的代表性图像。 所有图像均以相同的放大率拍摄。
(D-F图)用链霉素处理C57BL / 6小鼠或模拟处理。收集链霉素处理小鼠的器官后1天,进行分析,或者用指定的鼠伤寒沙门氏菌菌株混合物模拟感染或感染小鼠。在指定的时间点测定结肠内容物中丁酸(D),乙酸(E)和丙酸(F)的浓度。
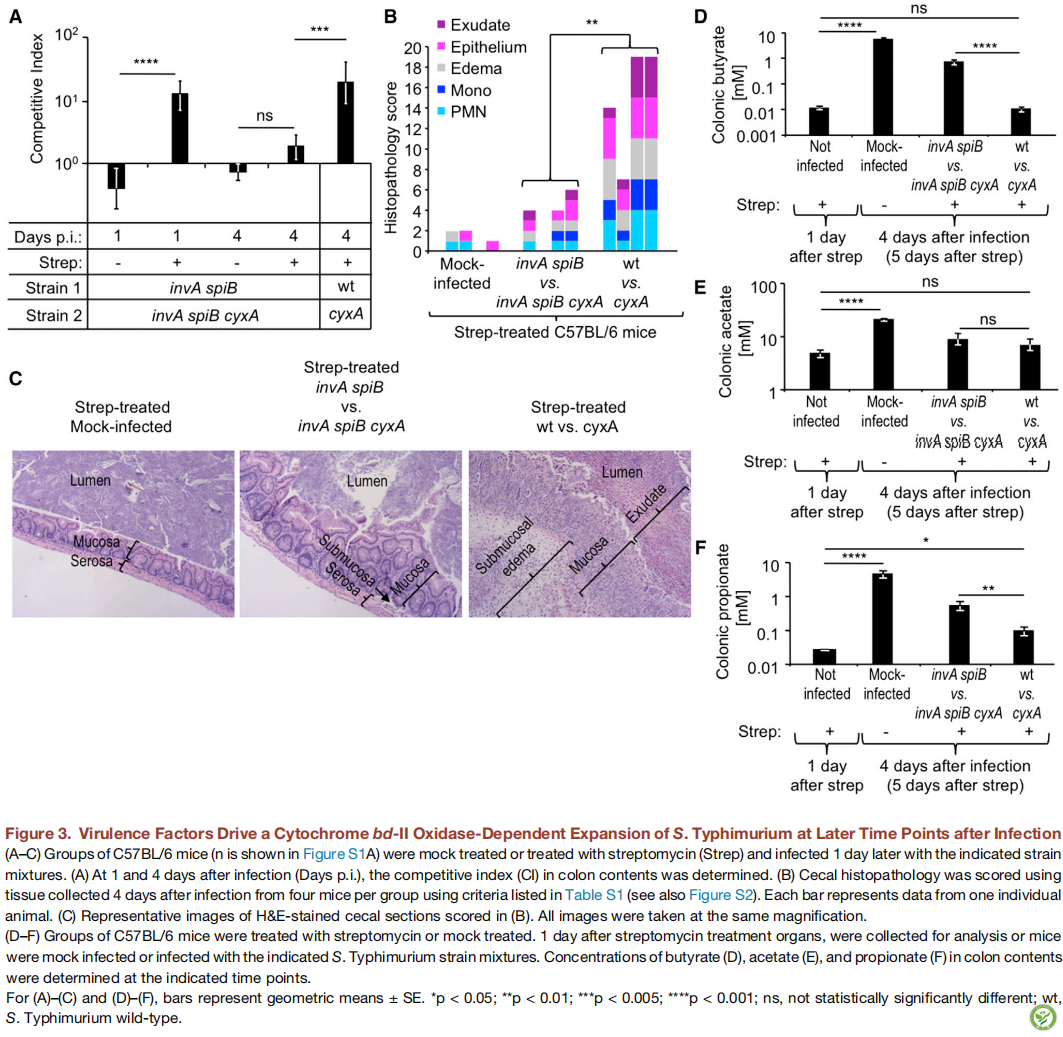
图4:鼠伤寒沙门氏菌的毒力因子诱导肠道炎症,并且在没有抗生素治疗的情况下使梭菌丰度减少
将CBA小鼠组(n = 6)模拟感染或用1×103 CFU /动物的毒性鼠伤寒沙门氏菌野生型(wt)或无毒的invA spiB突变体(invA spiB)胃内感染。
(A图)通过ELISA测定粪便脂质运载蛋白-2水平(也参见图S3)。
(B图)随着时间的推移在粪便中测定鼠伤寒沙门氏菌CFU
(C图)通过实时定量PCR确定梭菌16S rRNA基因复制数。
对于A-C图,数据点表示均值±标准误
(D图)在感染后第10天通过结肠内容物中存在的微生物群落的16S分析确定的种群系统发育组的平均相对丰度
(E图)感染后指定天数(天数p.i.)结肠内容物梭菌属成员的标准化丰度。
(F图)16S数据的加权主坐标分析,其中所有被鉴定为肠杆菌科的读数被排除在分析之外。有关16S数据的其他分析,请参见图S4-S7。
每个点代表一只动物的数据。* p <0.05; ** p <0.01; **** p <0.001
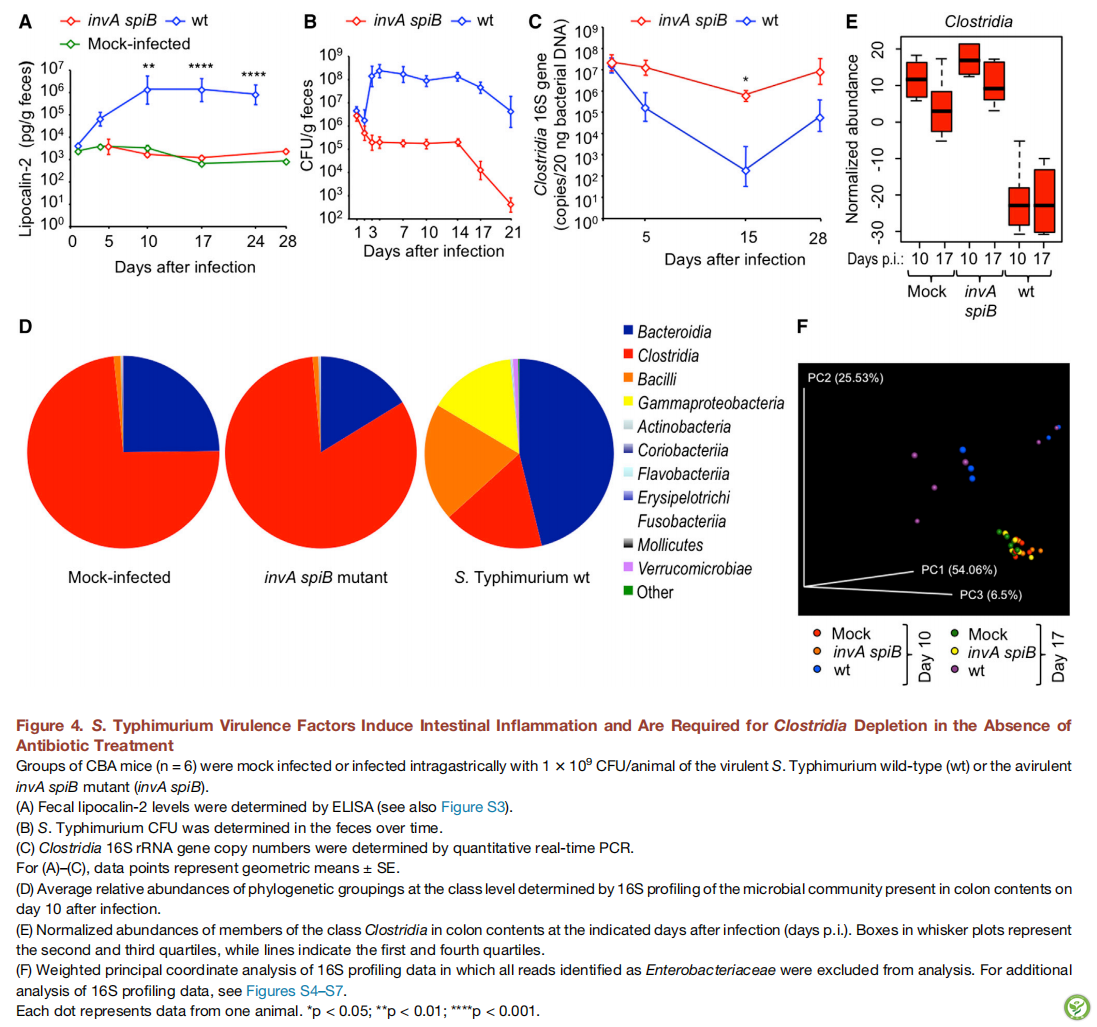
图5:细胞色素bd-II氧化酶和硝酸还原酶在没有抗生素治疗的情况下有助于肠腔内鼠伤寒沙门氏菌的扩增
(A图)用鼠伤寒沙门氏菌野生型(黑色圆圈)和呼吸缺陷型napA narZ narG cyxA突变体(蓝色圆圈)的1:1混合物胃内感染CBA小鼠组,感染后几天(天数pi)并在指定的时间收集样品。虚线红线连接从同一动物中回收的菌株。CI,竞争指数; **** p <0.001; ns:在统计上没有显著差异; nd:没有确定。
(B图)用1×103 CFU /动物的一种指定的鼠伤寒沙门氏菌菌株胃内感染CBA小鼠组(n = 4),并在感染后的指定天数收集样品。
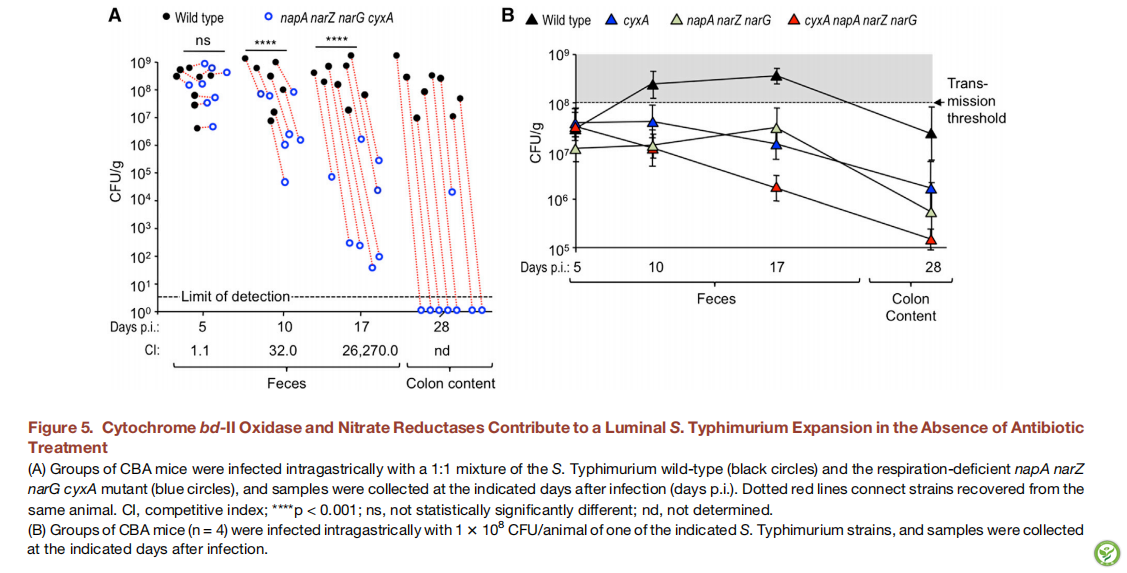
图6:梭菌的减少增加了结肠细胞生氧能力并促进依赖细胞色素bd-II氧化酶的鼠伤寒沙门氏菌的扩增
将一组CBA小鼠模拟感染或用鼠伤寒沙门氏菌野生型(wt)和cyxA突变体的1:1混合物胃内感染。在感染后第5,7和10天,将小鼠模拟接种或接种17种人源性梭菌分离物(C17)
(A图)在指定的时间点确定粪便或结肠内容物中的竞争指数(CI)。
A.D图)显示了盲肠内容物中丁酸(B),乙酸(C)和丙酸(D)的浓度。
对于(A)-(D),条形表示几何平均值±SE。
G图)在用DAPI核染色剂(蓝色荧光)复染的结肠切片中检测到哌莫硝唑(红色荧光)的结合。
以下显示了代表性的图像。 * p <0.05; *** p <0.005; **** p <0.001; ns,在统计学上没有显著性差异。
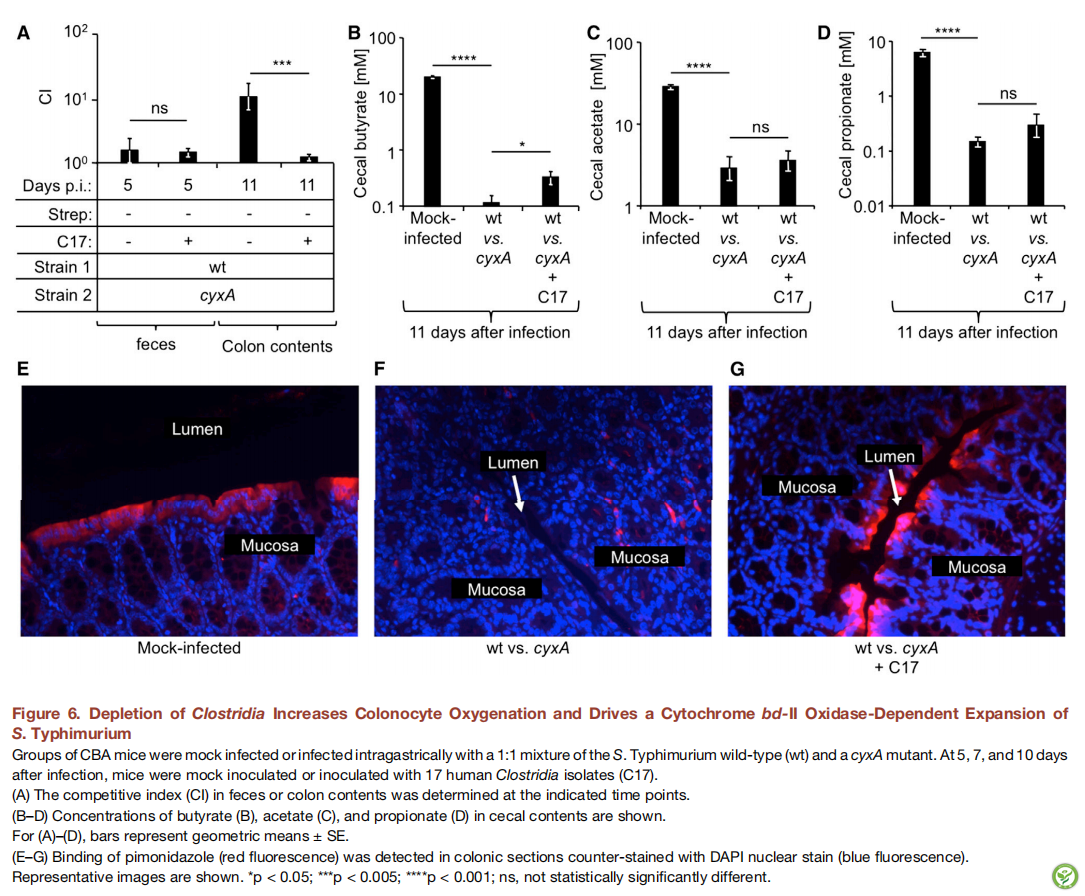
图7:鼠伤寒沙门氏菌通过呼吸可进行粪-口传播
(A和B图)用1×103CFU /动物的鼠伤寒沙门氏菌野生型,cyxA突变体或napA narZ narG cyxA突变体胃内感染CBA小鼠组(n = 6)。
感染后10天,将两只感染的小鼠(供体,黑圈)与两只幼稚小鼠(受体,红色)共同饲养。每笼一圈,在7(A)或18(B)后收集粪便(A)或结肠内容物(B)
Conclusion
沙门氏菌毒力因子和抗生素促进病原体扩增的机制相同,即:通过减少产丁酸梭状芽胞杆菌的数量,从而促进上皮细胞生氧,最终加速需氧沙门氏菌的繁殖。
Abstract
The mammalian intestine is host to a microbial community that prevents pathogen expansion through unknown mechanisms, while antibiotic treatment can increase susceptibility to enteric pathogens.Here we show that streptomycin treatment depleted commensal, butyrate-producing Clostridia from the mouse intestinal lumen, leading to decreased butyrate levels, increased epithelial oxygenation, and Salmonella restriction could be restored by tributyrin treatment. Clostridia depletion and aerobic Salmonella expansion were also observed in the absence of streptomycin treatment in genetically resistant mice but proceeded with slower kinetics and required the presence of functional Salmonella type III secretion systems. The Salmonella cytochrome bd-II oxidase synergized with nitrate reductases to drive luminal expansion, and both were required for fecal-oral transmission. We conclude that Salmonella virulence factors and antibiotic treatment promote pathogen expansion through the same mechanism: depletion of butyrate-producing Clostridia to elevate epithelial oxygenation, allowing aerobic Salmonella growth.
Conclusion
Salmonella virulence factors and antibiotic treatment promote pathogen expansion through the same mechanism: depletion of butyrate-producing Clostridia to elevate epithelial oxygenation, allowing aerobic Salmonella growth.
如您需原文,请联系本文作者和出版方,或请垂询肠动力研究院。本网站发布的所有资料将尽最大可能注明出处、作者及日期,如无意中侵犯了您的知识产权,请来信及时告知,我们将立即予以删除。
All information released by the WeChat Official Account will do its best to indicate the source, author and date. If we inadvertently infringe on your intellectual property, please inform us in time and we will delete it immediately.
 电话: 010-80854808
电话: 010-80854808
 传真:010-89525360
传真:010-89525360
 邮箱:dysbio@163.com
邮箱:dysbio@163.com
